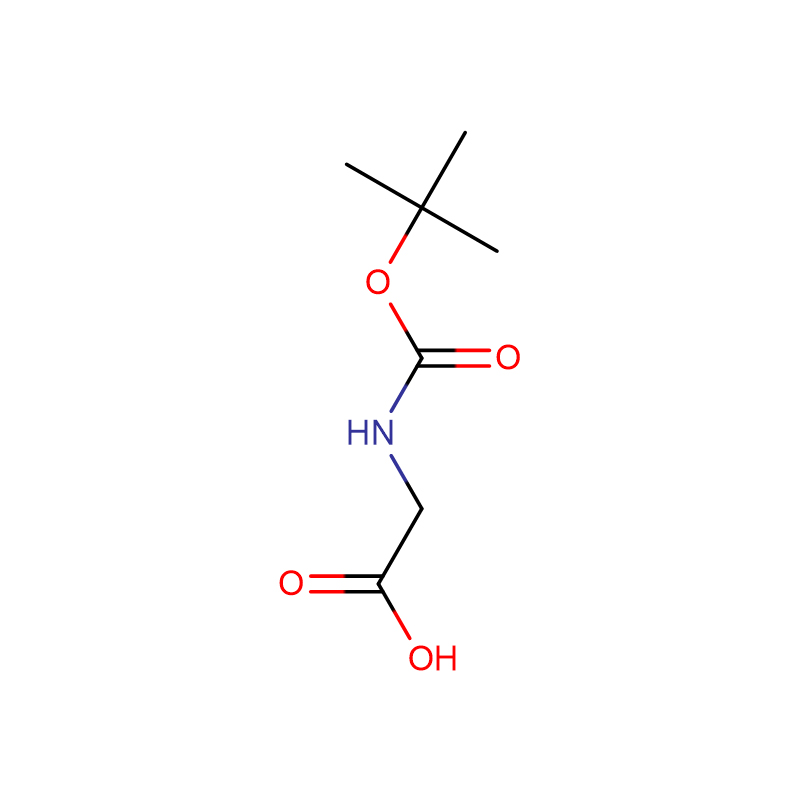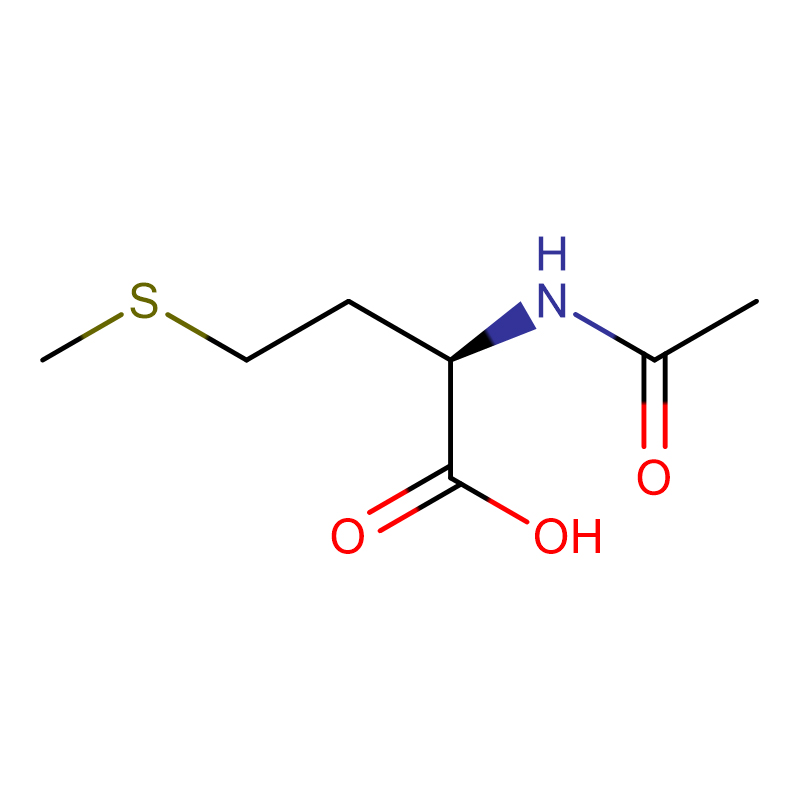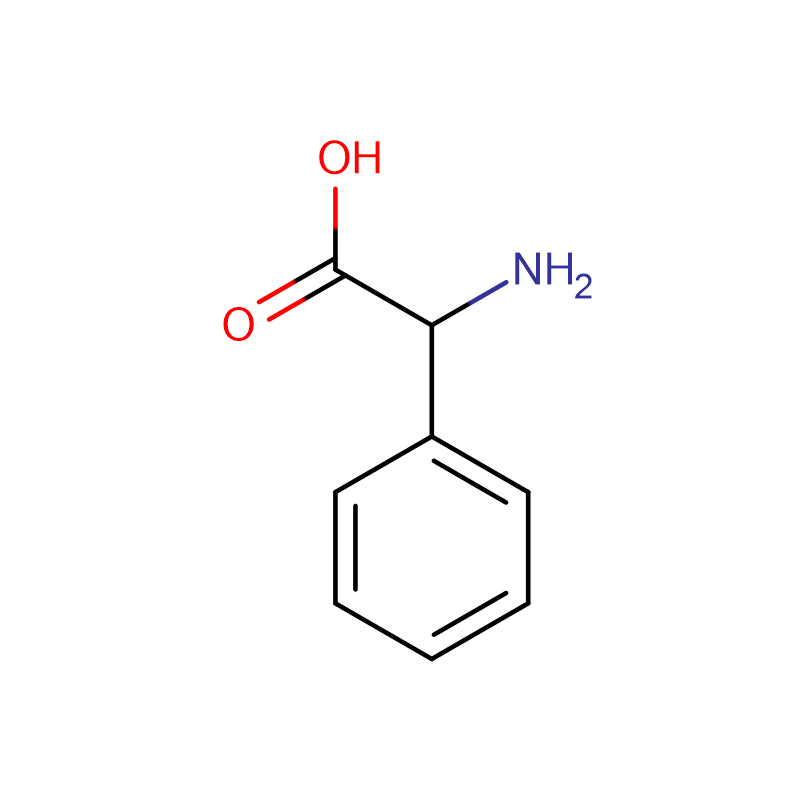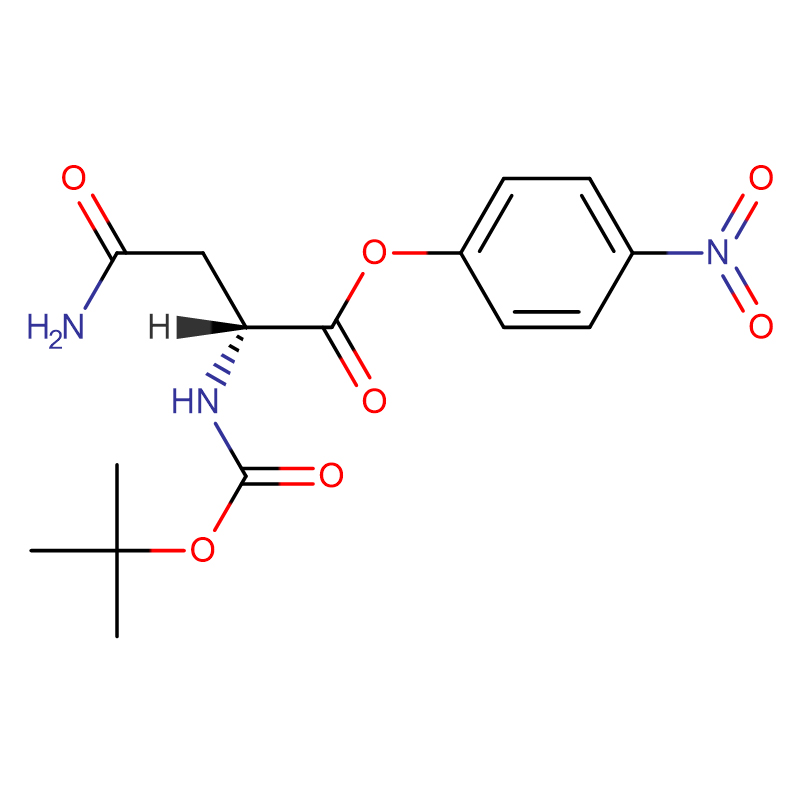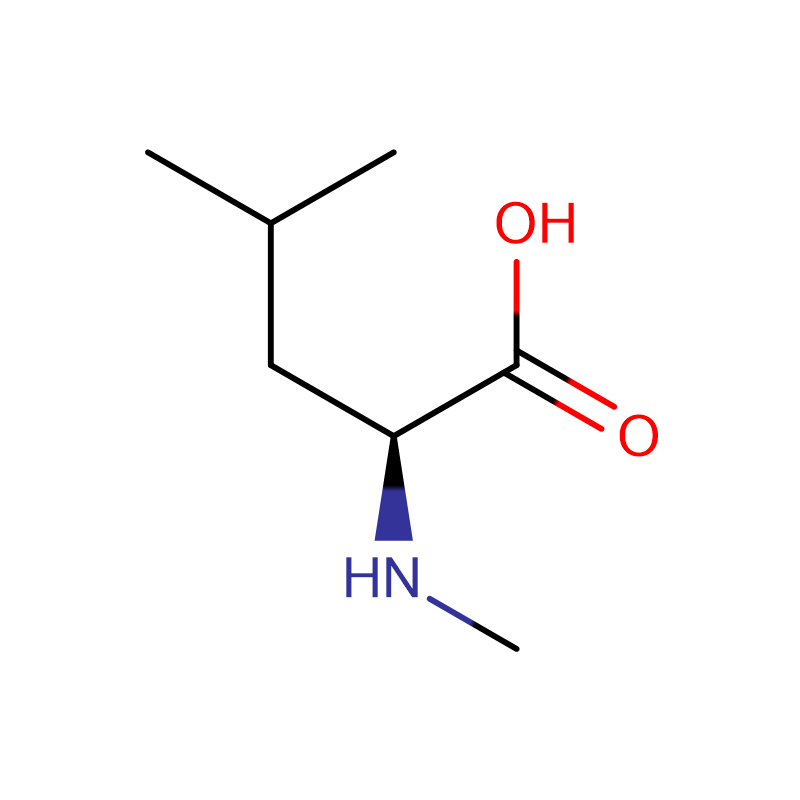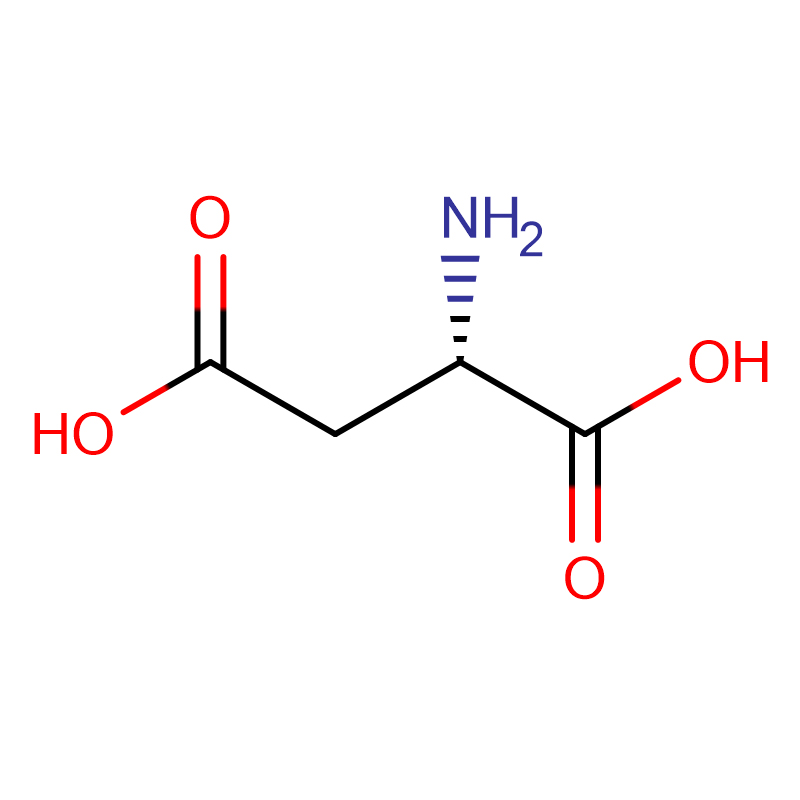
Л-аспарагинска киселина ЦАС:56-84-8 99% Бели прах
| Каталошки број | КСД90315 |
| Назив производа | Д-аспарагинска киселина |
| ЦАС | 56-84-8 |
| Молецулар Формула | Ц4Х7НО4 |
| Молекуларна тежина | 133.10 |
| Детаљи о складиштењу | 2 до 8 °Ц |
| Хармонизовани тарифни код | 29224985 |
Спецификација производа
| Изглед | Бели прах |
| Анализа | 99% |
| Оцена | УСП34 |
| Специфична ротација | +24,5 до +26 |
| Олово | <0,0005% |
| Губитак од сушења | <0,25% |
| Остатак при паљењу | <0,1% |
ЕААТ2 глутаматни транспортер, чини >90% преузимања глутамата хипокампуса.Иако је ЕААТ2 претежно експримиран у астроцитима, ∼10% ЕААТ2 молекула се налази у терминалима аксона.Упркос нижем нивоу експресије ЕААТ2 у глутаматергијским терминалима, када се резови хипокампуса инкубирају са ниском концентрацијом д-аспартата (ЕААТ2 супстрат), терминали аксона акумулирају д-аспартат једнако брзо као астроглија.Ово имплицира необјашњиву неусклађеност између дистрибуције ЕААТ2 протеина и транспортне активности посредоване ЕААТ2.Једна хипотеза је да је (1) хетероразмена унутрашњег супстрата са спољним супстратом знатно бржа од нето апсорпције и (2) терминали фаворизују хетероразмену због високих нивоа унутрашњег глутамата.Међутим, тренутно је непознато да ли хетероразмена и узимање имају сличне или различите стопе.Да бисмо решили овај проблем, користили смо реконституисани систем да упоредимо релативне стопе два процеса код пацова и мишева.Нето апсорпција је била осетљива на промене у мембранском потенцијалу и била је стимулисана спољашњим пермеабилним ањонима у складу са постојањем нераздвојене ањонске проводљивости.Коришћењем последњег, такође показујемо да брзина хетероразмене такође зависи од мембранског потенцијала.Поред тога, наши подаци даље указују на присуство цурења натријума у ЕААТ2.Укључујући нова открића у наш претходни модел узимања глутамата од стране ЕААТ2, предвиђамо да је осетљивост на напон размене узрокована трећим везивањем На(+) зависним од напона.Даље, и наши експерименти и симулације сугеришу да су релативне стопе нето узимања и хетероразмене упоредиве у ЕААТ2.